![]()
—— 中新系全媒体矩阵 ——
健康医疗行业垂直网站![]()
—— 中新系全媒体矩阵 ——
健康医疗行业垂直网站
大脑是哺乳动物中结构和功能最复杂的器官之一,其解剖区域形态独特,具有复杂的大脑网络连接、种类繁多的细胞类型,并且在大脑发育和衰老过程中存在高度整合的分子程序。大脑功能的实现需要一系列从神经递质到特定的复杂脂质(通常富含高度不饱和脂肪酸(FAs))的独特小分子化合物。目前,对于大脑代谢组的研究尚不充分,一是只专注于少数代谢物,二是研究设计和数据采集存在局限性,不能完整覆盖神经化学变化和全部大脑解剖区域。另外,由于反馈机制和调节途径众多,也不能简单地通过基因组或蛋白质组学特征预测大脑代谢物谱。大脑是身体的指挥中心和控制中心,至关重要,一旦出问题更加难以治。因此,对大脑的代谢组变化研究十分必要。本研究以小鼠为研究模型,通过非靶向代谢组学GC-MS和LC-MS平台对从青春期到老年阶段不同性别的小鼠10个大脑区域样本进行代谢组学分析,共鉴定1547种代谢物,发现大脑衰老过程中、大脑各区域之间存在显著代谢差异,而性别对大脑代谢物影响不大,为研究大脑在衰老过程中的时空代谢变化提供了宝贵信息,加速了我们对大脑的了解,并为研究一些与衰老相关的大脑功能障碍提供了基础。
部分研究结果
1 试验设计和不同年龄段10个大脑区域的代谢组分析此研究选取4组/每组8只雄性和8只雌性野生型(WT)小鼠,分别收集青春期(AD,3周龄)、成年早期(EA,16周龄)、中年(MA,59周龄)和老年(OA,92周龄)4个年龄段大脑的10个解剖区域:大脑皮层(CT)、嗅球(OB)、海马(HC)、下丘脑(HT)、基底节(BG)、丘脑(TL)、中脑(MB)、脑桥(PO)、延髓(MD)和小脑(CB),共640个样本(图1a),采用非靶向代谢组GC-MS和LC-MS平台进行分析,共鉴定到1547种代谢物,并使用ClassyFire分类系统将代谢物分为八类(图1b)。其中复合脂类代谢物占比最大(1103种,71.3%),这可能是因为大脑中内源性脂质含量较高,包括磷脂酰胆碱(PC)、磷脂酰乙醇胺(PEs)、三酰基甘油(TGs)、脂肪酸(FAs)、磷脂酰丝氨酸(PS)、磷脂酰肌醇(PIs)、鞘磷脂(SM)、神经酰胺(Cers)和二酰基甘油(DGs)等。有机酸类代谢物(包括氨基酸、修饰氨基酸、肽和qiang基酸)约占14%,剩余的15%主要包括有机氧化合物、有机杂环化合物、苯类化合物、有机氮化合物、核苷、核苷酸等。绝大多数代谢物广泛分布于所有10个大脑区域,以维持基本的大脑功能(图1c)。对代谢物进行通路分析,共发现118条相关代谢途径,其中前10条最重要的通路见图1d,表明通路分析的广度足以解释衰老过程中大脑区域的代谢变化。
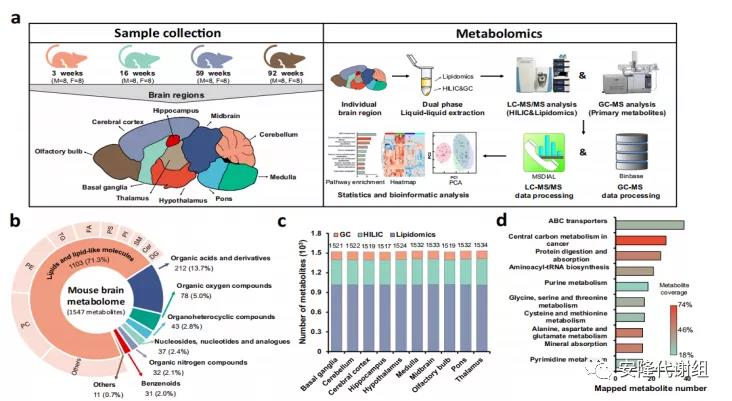
图1 试验设计和小鼠大脑代谢组
2 小鼠大脑区域代谢组的结构及年龄和性别的影响主成分分析(PCA)表明大脑区域和年龄对大脑代谢物的影响较大,几乎所有代谢物在大脑不同区域或年龄组之间都有很大的不同,而性别对大脑代谢物的影响较小(图2)。对10个大脑区域的相关矩阵分析显示,在每个脑结构分支内的代谢组之间存在高度正相关,但在大脑、脑干和小脑分支之间存在负相关(图3c)。大脑和脑干代谢组之间的负相关反映了不同区域的功能差异,大脑具有最复杂的神经元网络来指导更高的认知功能,而脑干可以调节自主活动。
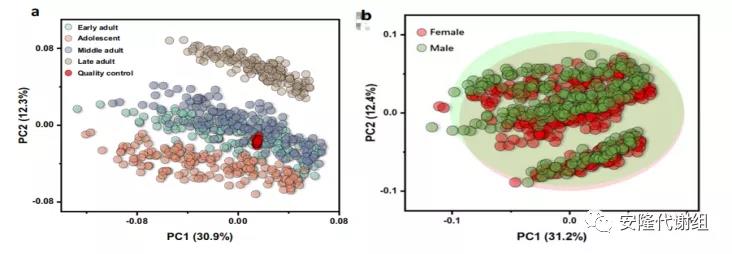
图2 年龄和性别对小鼠大脑代谢物的影响
研究表明,代谢物的变化分布在大脑的所有区域,并且存在明显的代谢异质性。与其它区域相比,脑干区域(PO和MD)中的脂质水平明显上调,而大脑区域(CT和OB)中许多脂质整体水平降低。脑干区域(MD和PO)中乙酰胆碱丰度最低,而CB组织在整个大脑中乙酰胆碱含量最低。这可能与乙酰胆碱可被乙酰胆碱酯酶(AChe)降解,AChe在MD、PO、MB中富集,在OB和CB中消耗有关。此外,大脑代谢组图谱显示BG中神经递质多巴胺及其代谢物的高度富集(图3e),腺苷及其类似物在BG中高度丰富,并且与腺苷A2a受体(Adora2a)和磷酸二酯酶10(PED10,一种cAMP水解酶)的原位杂交图像完全一致(图3f)。匹配高丰度的代谢物和基因表达有助于基因功能的验证。鸟嘌呤在CB中高度表达,并且与浦肯野细胞蛋白2(Pcp2)完全共定位(图3g),为Pcp2作为鸟嘌呤核苷酸解离抑制剂的功能提供了依据。
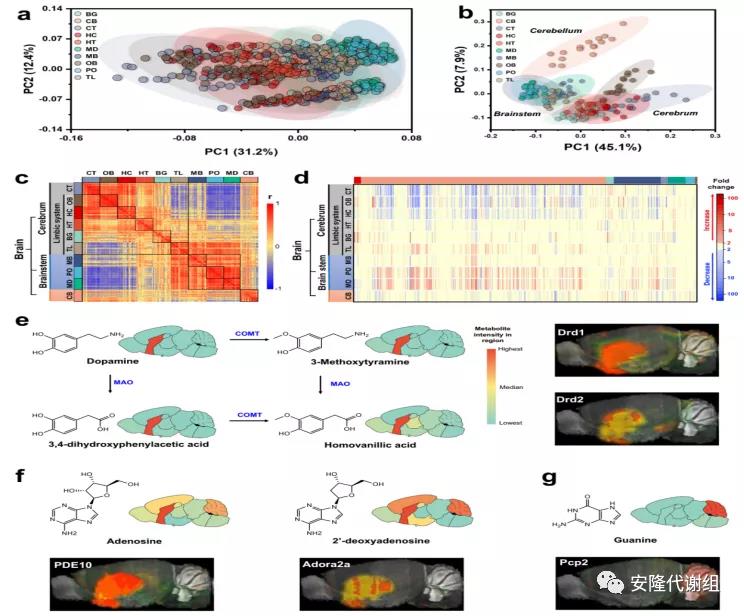
图3 小鼠大脑不同区域代谢差异
3 衰老对小鼠大脑代谢组的影响在衰老过程中,大脑会经历一系列结构和功能的变化。主成分分析(PCA)表明,不同年龄之间代谢组存在显着差异(图4b)。通过相关性分析发现,从青春期到成年早期,脑干和大脑代谢组从高正相关转变为高负相关,不同大脑区域的内部相关性显著减弱。在从成年早期到中年的衰老过程中,脑干和大脑区域之间的强负相关性也严重减弱,而在老年期时,几乎所有的负相关都消失了(图4a)。表明,大脑功能在成熟时期特异性增加,而在衰老过程中去分化。先前的研究表明,大脑发育和衰老以特定于区域的方式异步发生,而不是在所有区域均匀发生,在每个区域的代谢组相关热图中发现了类似的老化趋势(图4c)。
大脑和小脑代谢组的变化需要应对大脑发育和衰老过程中的重大变化。通过个体代谢物水平的变化也可以看出代谢调节的这些差异。从青春期到成年早期,37.1%的代谢物水平发生显著变化,从成年早期到中年期只有10.0%的代谢物发生显著变化,而从中年期到老年期,61.5%的代谢物发生了改变(图4d)。
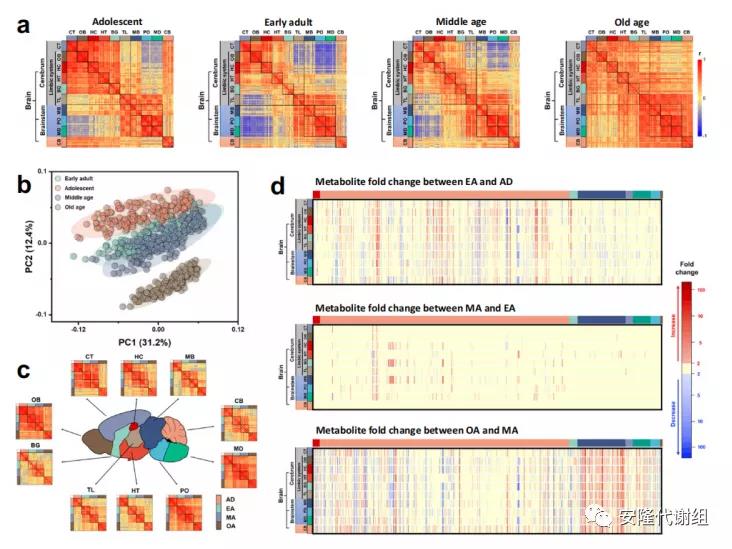
图4 衰老对小鼠大脑代谢组的影响
4 衰老小鼠大脑中的鞘脂动力学通过此研究,还发现了三种鞘脂(鞘磷脂(SMs)、己糖神经酰胺(HexCers)及其硫脂(sHexCers))随年龄的交替模式,表明小鼠大脑代谢组图谱可以深入了解生物过程。热图分析显示,SMs、HexCers和sHexCers从青春期到成年早期大幅增加,而从中年期到老年期大幅减少,尤其是具有C20-26脂肪酰基的HexCers和sHexCers(图5a)。这一发现与神经酰胺合酶2(CerS2,一种主要在少突胶质细胞中表达以产生C20-26 Cers的酶)在3周龄大脑中上调,在60周龄大脑中下调一致。有报道称,衰老的大脑和阿尔茨海默病患者的大脑都表现出CerS2活性的丧失,并伴有髓鞘变性,表明长酰基链对维持大脑髓鞘功能和完整性的重要性。此外,只有4种SM在中年和老年期之间存在显著变化,并且在区域之间、衰老期间和性别之间存在巨大差异。其他鞘脂在衰老过程中表现出减少,例如短链脂肪酰基衍生物,尤其是HexCer和sHexCer。图5c展示了从青春期到老年期髓鞘化演变过程。特定的酰基链长度对HexCer、sHexCer和SM有特别影响,对于长度超过18个碳的酰基(和长链SM)具有不同的模式,通常为单不饱和形式。这种结构可能支持成年或衰老大脑中的髓鞘可塑性。
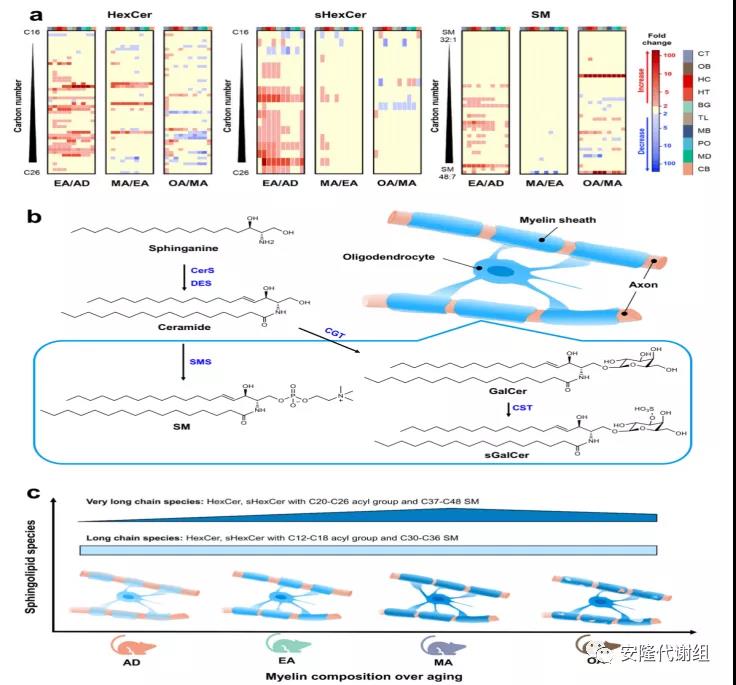
图5 小鼠大脑中鞘脂的动力学
延伸与讨论
综上所述,此研究以小鼠为研究模型,通过对不同年龄、性别的小鼠10个大脑区域样本进行代谢组学分析,共鉴定1547种代谢物,发现大脑衰老过程、大脑各区域之间存在显著代谢差异,而性别对大脑代谢物影响不大。大脑发育的分化-去分化轨迹表明每个区域都有特定的衰老程序,加速了我们对大脑的了解,并为研究痴呆症或其它与衰老相关的大脑功能障碍提供了方向。 参考文献:A metabolome atlas of the aging mouse brain,IF=14.9 杂志:Nature Communications年份:2021年封面图来源:文献原文